一种水生植物根系立体几何构型的构建方法
阅读说明:本技术 一种水生植物根系立体几何构型的构建方法 (Method for constructing three-dimensional geometric configuration of aquatic plant root system ) 是由 黄晓龙 李松阳 李宽意 于 2021-08-24 设计创作,主要内容包括:本发明公开了一种水生植物根系立体几何构型的构建方法,包括如下步骤:1)通过扫描仪获取高分辨率水生植物根系图像;2)对根系图像进行处理,得出根系一级侧根个数、一级侧根长度、二级侧根个数、二级侧根长度、二级侧根直径、顶端不分枝区域长度、末端不分枝区域长度、分支间距、分支数目、主根与根毛夹角和/或初始生长速度的结构数据;3)建立根系的几何构型。本发明实现了水生植物根系构型可视化,可以较为较真实地模拟水生植物根系的生长过程,为研究分析根系周围营养元素对水生植物根系生长发育的可塑性影响等提供了一种简单直观的工具,为水生植物保护、管理和生态修复等提供了科学依据。(The invention discloses a method for constructing a solid geometric configuration of a root system of an aquatic plant, which comprises the following steps: 1) acquiring a high-resolution aquatic plant root system image through a scanner; 2) processing the root system image to obtain structural data of the number of primary lateral roots, the length of the primary lateral roots, the number of secondary lateral roots, the length of the secondary lateral roots, the diameter of the secondary lateral roots, the length of a top non-branching region, the length of a tail end non-branching region, branch spacing, the number of branches, the included angle between a main root and root hairs and/or the initial growth speed of the root system; 3) the geometric configuration of the root system is established. The invention realizes the visualization of the configuration of the aquatic plant root system, can simulate the growth process of the aquatic plant root system more truly, provides a simple and intuitive tool for researching and analyzing the plasticity influence of nutrient elements around the root system on the growth and development of the aquatic plant root system, and the like, and provides scientific basis for the protection, management, ecological restoration and the like of the aquatic plant.)
技术领域
本发明属于植物仿真技术领域,具体涉及一种水生植物根系立体几何构型的构建方法。
背景技术
植物根系的演变是使植物从水生环境迁移到陆生环境的基础,并最终导致叶片合成糖类,根系供应资源(主要是水分和营养)之间功能上的分割。作为植物三大营养器官之一,根系在生物圈物质循环和能量流动中起到关键性作用。根系构型在植物资源利用中起到重要作用。
植物根系构型是指根系在生长介质中的形状和分布,包括同一根系的各种根在介质中沿根轴二维平面上的分布(平面几何构型)和不同类型的根系在介质中的三维空间分布,这两种构型都可以通过根系拓扑指数表现出来。近几十年对植物根系构型研究方法可分为两大类:
第一类方法是通过扫描或微根管法(Minirhizotron)获取植物真实根系构型,之后通过软件(WinRhizo、SimRoot)等分析根系形态和拓扑指标。这类方法获取数据较为真实,可获取根系连续生长数据,通过对根系二维构型建模可以反映根系三维立体几何构型。然而通过挖掘的方法获取植物根系工作量大,对于森林乔木根系只能通过局部挖掘来反映根系情况。通过植物根系二维构型来构建根系三维构型不可避免地会导致根系部分信息缺失。此外这种根系模型也较难适用于已经完成生长的根系。
第二类方法是通过构建植物形态和结构的模型对植物的形态结构进行量化研究,总结出植物生长的规律,根据植物生长情况对植物根系构型模型进行修正上,再用适当的方式对植物的形态结构进行量化表达,可以简化根系模型,减少计算量。第二类方法以近几十年发展出通过计算机模拟重现植物根系发育过程的L系统为代表,可以较好模拟植物生长状况。L系统由Aristid Lindenmayer于1968年提出并以他的名字首字母命名,之后被Przemyslaw Prusinkiewicz等人发展用于植物和植物器官的生长模拟。这一类方法对陆生植物根系构型的假设都建立在植物结构具有对称性(symmetry)和自相似性(self-similarity)两大前提条件上,也就是植物组织结构的几何形状在局部和整体上具有相似性假设上,应该说从这两个假设出发建立的根系构型模型能较好地符合实际情况,在植物叶镶嵌、茎分枝和花序的模拟结果较好。通过植物局部根系模拟整体根系特征,动态地对水稻等植物根系在多种营养条件下进行三维空间模型构建,可以较好地表征根系拓扑结构。然而目前有研究发现一种水生植物凤眼莲(Eichhornia crassipes)的根系能较好地满足对称性假设,却并不满足自相似性假设。凤眼莲局部根系与整体根系在结构上并不一致,其整体根系为由多条鱼骨状构型根系构成一种特殊的叉状分支构型,称作“辐射状聚合构型”。在另外一种水生植物大薸(Pistia stratiotes)中也发现类似的情况。因此在水生植物中建立根系模型,要慎重考虑根系特殊性,根据情况探讨自相似性在水生植物中的适用性。
目前植物根系的研究方法中,常用的微根管法能够非接触地获取根系结构信息,针对小麦、水稻和大豆作物已经进行了大量研究。然而微根管法只适用于陆生植物,水生植物根系埋藏于饱含水分的底质中,无法通过微根管法实时传输数据研究,因此之前对于水生植物根系研究更多地聚焦于对根系形态特征和功能性状,对根系结构关注相对较少,更缺乏通过测量根系生长指标建立动态生长模型的报道。
发明内容
本发明提供一种水生植物根系立体几何构型的构建方法,实现了水生植物根系构型可视化。
为解决上述技术问题,本发明所采用的技术方案如下:
一种水生植物根系立体几何构型的构建方法,包括顺序相接的如下步骤:
步骤(1):通过扫描仪获取1:1高分辨率(≥600dpi)水生植物根系图像;
步骤(2):使用根系测量分析软件对步骤(1)的根系图像进行处理,得出根系一级侧根个数、一级侧根长度、二级侧根个数、二级侧根长度、二级侧根直径、顶端不分枝区域长度、末端不分枝区域长度、分支间距、分支数目、主根与根毛夹角和/或初始生长速度等结构数据;
步骤(3):根据步骤(2)得到的根系结构数据,建立根系的几何构型。
L系统的核心概念是迭代置换(rewriting),即基于迭代置换规则,又称产生式(production)p在迭代过程中对简单的初始目标部分或全部不断的置换从而表达复杂对象的一种方法。L系统以字符为核心,以字符的产生式规则为置换规则。L系统内部用一个字符串表示系统的状态,称为L字符串。系统的初始状态,即初始L字符串称为公理。产生式p即是L字符串中字符的迭代置换规则。在每个演化周期,L系统应用产生式对系统状态(L字符串)进行修改,形成新的系统状态。因此L系统最后得到的是一个由特定字母组成的字符串,通过给L系统中每一个字母赋予一个特定的图形含义,从而实现L系统图形说明。考虑仅有两个字母a和b构成的系统ab,假定产生式p为a→ab和b→a,即a被ab置换,b被a置换。根据规则ab第一次被置换为aba,第二次被置换为abaab,第三次被置换为abaababa,第四次被置换为abaababaabaab,第五次被置换为abaababaabaababaababa……由于植物根系的生长可以看做是一个连续和重复的过程,因此通过L系统可以较好地模拟植物根系的动态生长过程。
目前在陆生植物中基于植物根系具有对称性和自相似性假设建立的L系统已经做了较好研究,但目前研究发现一些水生植物如凤眼莲(Eichhornia crassipes)的根系构型并不符合L系统自相似性假设,本申请通过对L系统的改进,构建了不符合L系统自相似性假设的水生植物根系的立体几何构型。
本申请通过模型构建,可以模拟水生植物从幼苗到成熟个体,从简单根系到复杂根系的动态生长过程。可从根系构型这一角度阐述水生植物对水生生态系统的适应性,也可为水生植物保护、管理和生态修复提供科学依据。
上述步骤(3)中,根系的几何构型包括根系伸长规则和根系分支规则。
上述根系伸长规则包括根系长度计算、根伸长规则和初始根系生长方向。
根系长度计算:当单根系根龄小于等于该单根系根寿命(rlt)时,该单根系会一直伸长,根系长度通过公式(1)计算:
参数lmax表示该单根系最大长度,r表示该单根系初始伸长速度,t为该单根系根龄;l(t)表示该单根系在t时间的长度;
根伸长规则:单根系会朝着一定的方向伸长,根伸长规则通过公式(2)描述:
符号G代表根生长规则,表示根在当前长度为l,当前时刻为t的条件下,经过生长间隔Δt后的生长变化,参数l表示通过公式(1)计算的t时间的根长;Δt为根系的生长时间间隔;Δl为经过Δt时间的根长增量;R表示根系生长方向;符号FΔl表示长度为Δl的根段,通过公式(2)的重复迭代能够完成根系朝着方向R长度为Δl生长;
初始根系生长方向:初始根系生长方向R由主根与侧根的夹角θ和径向角γ确定,其中夹角θ是从对真实植物根系计算得到的均值和标准差的正态分布中随机取值;径向角γ表示侧根围绕主根旋转的角度,从0到2π的均匀分布中随机取值(图1所示)。在根系生长过程中,为使得生成的根系更加具有随机性,从而更符合植物生长实际情况,对初始根系生长方向增加随机角度变化,从0到2π的均匀分布中随机取值,即长度为Δl的根段生长时,设置根段生长尺度dx(dx<Δl),随后依据增加随机角度变化的生长方向增长dx长度,直到完成Δl长度生长为止。
根系分支规则如下:分枝级数为i的单根系能够产生分枝级数为i+1的侧根,单根系分为三个区域:顶端不分枝区域(la)、末端不分枝区域(lb)以及分支区域(ln*N)(图2所示),ln为分枝级数为i的单根系分支间距,N表示分枝级数为i的单根系最大分支数目;
当单根系长度大于顶端不分枝区域和末端不分枝区域长度之和时,会产生分支,单根系产生第j个分支时的根长通过公式(3)计算:
lj=la+lb+(j-1)*ln 公式(3)
结合公式(1)可以计算出单根系产生第j个分支时的延迟时间dj为:
根系分支规则通过公式(5)和(6)描述:
B(j)->{D(0,dj)j≤N 公式(5)
其中,B表示根分支产生的规则,参数j表示产生的分支为第j个,B(j)代表了第j个分支的产生规则,D表示延迟规则,d是第j个分支的延迟时间,可以通过公式(4)计算,D(0,dj)表示第j个分支经过延时时间dj后,该分支会产生:
其中D表示t时间的分支数目,其中tend为分支结束时间;Δt为根系的生长时间间隔,公式(6)解释了延迟规则具体的符号替代过程,其中,tend采用公式(5)替代,tend替代后即为dj,当前时间t经过生长时间间隔Δt后,如果大于dj或者tend,就会产生新分支B,记为Bnew.该分支Bnew是B的第j+1个分支,并且Bnew开始调用生长规则G开始生长,B(1)表示Bnew会产生第1个分支。
作为一种具体的实现方案,步骤(1):通过扫描仪获取一条根系1:1高分辨率(≥600dpi)水生植物根系图像;步骤(3):根据步骤(2)得到的根系结构数据,建立一条根系的几何构型,再推广为整株根系的几何构型。
上述方法,剪切其单独根系分别扫描,再组合成根系构型的方法来研究其真实的整体根系结构;或者,建立一条根系的几何构型后,根据根系之间的规律,推广为整株根系的几何构型。
上述步骤(3)中,建立一条根系的几何构型后,重复步骤1)-2)建立其余根系的几何构型,再组合成整株根系的几何构型;或者,建立一条根系的几何构型后,根据研究获得的真实根系之间的规律,推广为整株根系的几何构型。
如构建凤眼莲的根系立体几何构型时,首先建立凤眼莲单独一条根系的模型,通过对局部根系立体结构较为真实的模拟,再组合或推广成整个植株的结构,从而较真实地模拟凤眼莲等水生植物根系的生长过程。
本发明未提及的技术均参照现有技术。
本发明一种水生植物根系立体几何构型的构建方法,实现了水生植物根系构型可视化,可以较为较真实地模拟水生植物根系的生长过程,为研究分析根系周围营养元素对水生植物根系生长发育的可塑性影响等提供了一种简单直观的工具,为水生植物保护、管理和生态修复等提供了科学依据。
附图说明
图1为本发明水生植物根系生长方向图;
图2为本发明水生植物根系分支自相似结构;
图3为本发明水生植物根系可视化过程;
图4为凤眼莲整体根系构建视图,其中,4A为实物图,4B为构建图;
图5为凤眼莲单独一条一级根系构建视图,其中,5A为单独一条一级根系的模型,5B为单独一条根系的模型。
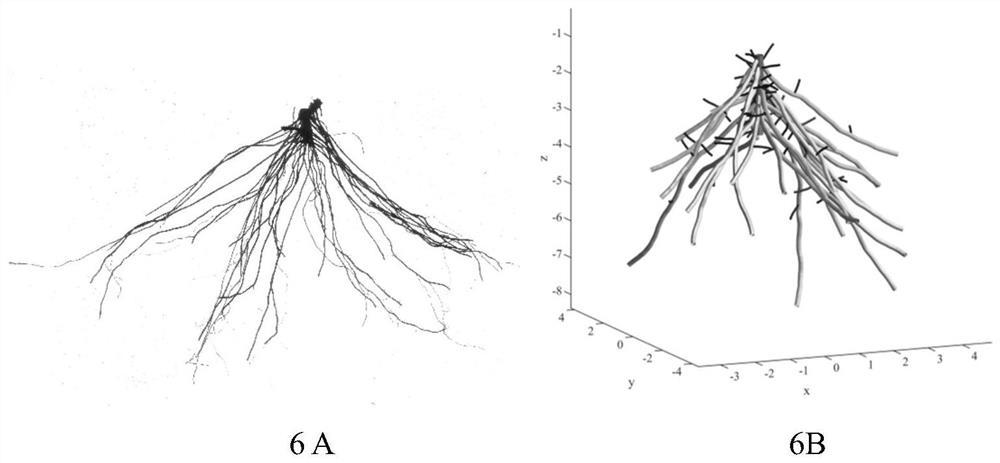
图6为穗花狐尾藻整体根系构建视图,其中,6A为实物图,6B为构建图;
图中,1为主根,2为侧根,3为分支级数i的单根,4为分支级数i+1的侧根,5为根毛。
具体实施方式
为了更好地理解本发明,下面结合实施例进一步阐明本发明的内容,但本发明的内容不仅仅局限于下面的实施例。
实施例1
外来入侵植物凤眼莲又名水葫芦、凤眼蓝,属于水生植物中的漂浮植物,原产于南美洲亚马逊河流域。凤眼莲原作为观赏植物引入中国,逸生后在长江流域和南方河流、湖泊等生境水面形成连绵成片漂浮植毡层,遮蔽阳光,阻隔水体和大气交换,引起沉水植物、鱼类和软体动物等生物死亡,已列入IUCN《100种世界最严重外来入侵物种名录》和第一批《中国外来入侵物种名单(2003)》。凤眼莲的根系非常复杂,重叠很多,常规扫描分析方法无法得到其完整根系。通过剪切其单独根系分别扫描,再组合成根系构型的方法来研究其真实的整体根系结构。
在实验室中培养凤眼莲植株。将凤眼莲根茎连接处剪断后,根系用镊子放置在扫描仪上透明水槽中,加自来水至水槽容积的2/3左右。用镊子展开根系,使根系处于分散无重叠状态。一般一次只扫描一条根系;若根系较小,则一次可扫描多条根系。根系图像使用爱普生12000XL扫描仪(Seiko Epson Corp.,Suwa,Nagano,Japan)扫描获取。图像分辨率设置为600dpi,扫描完成后保存1:1无压缩8bit高分辨率tiff格式图像。此扫描仪配备有两种光源:白色LED光源和红外IR光源。本发明使用WinRHIZO根系测量分析软件(RegentInstruments Inc.,Québec City,Canada)对扫描仪扫描后根系图像进行处理,此软件利用非统计学方法分析根系图像信息,得出根系结构数据。为使根系与背景对照明显,对于接近透明的凤眼莲根系预先使用0.2mmol·L-1(0.075mg·mL-1)甲烯蓝溶液染色30s后再使用白色LED光源扫描,背景选用扫描仪内侧配套的白色背景板。凤眼莲根系较为独特,它可以看作所有一级根系都生长在一个生长点上(图4A)。生长末期的凤眼莲的一级根系数量超过100个,每一个单独的一级根系都有且只有一条主根,在主根上密生非常多的二级根系(图4A)。基于凤眼莲单独一条一级根系实际生长状况,设置在根系生长周期内,每间隔3天产生一个分支级数0的主根,可以建立凤眼莲根系的模型。因此首先建立凤眼莲单独一条一级根系的模型(图5A)。通过对单独一条一级根系立体结构较为真实的模拟,再组合成整个植株根系的几何构型,从而较真实地模拟凤眼莲生长过程。凤眼莲最少只需12天即可扩张一倍的种群,在本例中对凤眼莲进行30天的生长模拟,在实践中这段时间足够凤眼莲完成生长过程。
凤眼莲单独一条一级根系的构成主要由分支级数为0的主根和密生的分支级数为1的根毛构成,其中根毛不再产生分支(图5A)。
测量凤眼莲根系单根长度、分支间距、顶端不分枝区域、末端不分枝区域和分支数目等参数(表1),可以建立凤眼莲单独一条根系的模型(图5B)。
整个植株根系的几何构型包括根系伸长规则和根系分支规则;
根系伸长规则包括根系长度计算、根伸长规则和初始根系生长方向;
根系长度计算:当单根系根龄小于等于该单根系根寿命(rlt)时,该单根系会一直伸长,根系长度通过公式(1)计算:
参数lmax表示该单根系最大长度,r表示该单根系初始伸长速度,t为该单根系根龄;l(t)表示该单根系在t时间的长度;
根伸长规则:单根系会朝着一定的方向伸长,根伸长规则通过公式(2)描述:
符号G代表根生长规则,表示根在当前长度为l,当前时刻为t的条件下,经过生长间隔Δt后的生长变化,参数l表示通过公式(1)计算的t时间的根长;Δt为根系的生长时间间隔,本研究中设为1天;Δl为经过Δt时间的根长增量;R表示根系生长方向;符号FΔl表示长度为Δl的根段,通过公式(2)的重复迭代能够完成根系朝着方向R长度为Δl生长;
初始根系生长方向:初始根系生长方向R由主根与侧根的夹角θ和径向角γ确定,其中夹角θ是从对真实植物根系计算得到的均值和标准差的正态分布中随机取值;径向角γ表示侧根围绕主根旋转的角度,从0到2π的均匀分布中随机取值(图1所示);在根系生长过程中,为使得生成的根系更加具有随机性,从而更符合植物生长实际情况,对初始根系生长方向增加随机角度变化,从0到2π的均匀分布中随机取值,即长度为Δl的根段生长时,设置根段生长尺度dx(dx<Δl),随后依据增加随机角度变化的生长方向增长dx长度,直到完成Δl长度生长为止。
根系分支规则:
分枝级数为i的单根系能够产生分枝级数为i+1的侧根,单根系分为三个区域:顶端不分枝区域(la)、末端不分枝区域(lb)以及分支区域(ln*N)(图2所示),ln为分枝级数为i的单根系分支间距,N表示分枝级数为i的单根系最大分支数目;
当单根系长度大于顶端不分枝区域和末端不分枝区域长度之和时,会产生分支,单根系产生第j个分支时的根长通过公式(3)计算:
lj=la+lb+(j-1)*ln 公式(3)
结合公式(1)可以计算出单根系产生第j个分支时的延迟时间dj为:
根系分支规则通过公式(5)和(6)描述:
B(j)->{D(0,dj) j≤N 公式(5)
其中,B表示根分支产生的规则,参数j表示产生的分支为第j个,B(j)代表了第j个分支的产生规则,D表示延迟规则,d是第j个分支的延迟时间,可以通过公式(4)计算,D(0,dj)表示第j个分支经过延时时间dj后,该分支会产生:
公式(6)解释了延迟规则具体的符号替代过程,其中,tend采用公式(5)替代,tend替代后即为dj,当前时间t经过生长时间间隔Δt后,如果大于dj或者tend,就会产生新分支B,记为Bnew.该分支Bnew是B的第j+1个分支,并且Bnew开始调用生长规则G开始生长,B(1)表示Bnew会产生第1个分支。
采用根系伸长规则G和根系分支规则B植物即可以进行生长和分支,设置初始状态为GB(j),其中j值可以设置为1,可以表示单根或者初始分支,假设根系初始状态为GB(1),过程GB(1)->RFGD->RFRFGD->RFRFRFGB(j+1)GB(1)展示了经过三次此迭代后的字符串,该过程可视化过程如图3所示,其中可视化过程中G、D、B、R等符号没有对应的可视化图形,F对应某种可视化图形(比如线段)。GB(1)表示一个新的种子或者初始分支,经过第一次生长,GB(1)变为RFGD,可视化时依据符号R包含的夹角θ和径向角γ取值,从而可视化展示根段F.经过三次生长,GB(1)变为RFRFRFGB(j+1)GB(1),其中RFRFRF表示该单根或者分支有3个根段,GB(j+1)表示该单根会继续生长,并产生第j+1个分支。
通过上述几何构型的构建,可以计算出在一个成熟的凤眼莲根系总根毛长度的2,022.54m(根毛长度×根毛个数a×一级根系个数)(表2),加上凤眼莲一级根系主根的长度20.24m(主根长度×一级根系个数c),一株凤眼莲成熟根系的总长度可达到惊人的2,042.78m。上述几何构型较真实地模拟了凤眼莲根系的生长过程,与凤眼莲的实际生长吻合。
表1凤眼莲单独一条一级根系模型参数
表2凤眼莲整株初生和成熟根系指标
实施例2
穗花狐尾藻(Myriophyllum spicatum)又名穗状狐尾藻,在我国南北各地均有,生于池塘或河川中,为世界广布种。穗花狐尾藻的根系为典型的叉状分支构型,由于大多数水生植物根系都是叉状分支构型,因此选择这种植物根系进行生长模拟构建具有代表性。
根系图像使用爱普生12000XL扫描仪(Seiko Epson Corp.,Suwa,Nagano,Japan)扫描获取。对于浅色的穗花狐尾藻根系使用红外光源,背景为扫描仪内侧配套的无色玻璃。扫描完成后取出根系,倒出水槽中剩余水分,用自来水冲洗清洁水槽内附着的残留根系后,再用同样的方式扫描下一条根系。
穗花狐尾藻的根系有以下特征:具有健壮发达的一级根系,而每一个一级根系上二级根系不发达(如图6A)。其幼苗根系平均有26.2个一级根系,平均长度为32.31mm(表3)。幼苗根系的一级侧根平均有3个分支,分支间平均距离为10.77mm。成熟穗花狐尾藻根系的二级根系也是不多的,11个左右,平均距离12.09mm。据此可以计算出穗花狐尾藻成熟根系的总长度为73.08m(一级侧根长度×一级侧根个数c+二级侧根长度×二级侧根个数a×一级侧根个数)。
依据获取的穗花狐尾藻根系生长数据,设置基于L系统的穗花狐尾藻根系模型参数(表4),建立L系统根系立体几何构型(具体同实施例1),从而较真实地模拟穗花狐尾藻生长过程(图6B),与穗花狐尾藻的实际生长吻合。对穗花狐尾藻进行了90天的生长模拟,在实践中足够穗花狐尾藻完成生长过程。
表3穗花狐尾藻整株初生和成熟根系指标
表4穗花狐尾藻根系指标
- 上一篇:一种医用注射器针头装配设备
- 下一篇:一种红薯水培育苗方法